The bacteria (E. coli) has been the prime
experimental organism for studies on homologous recombination, and
investigations of meiosis have mostly been performed using lower
eukaryotes such as the baker's yeast (Saccharomyces cerevisiae),
and fruit flies (Drosophila). More recent work with mutated mice
has also provided information on these reactions in mammals. This
research has shown that substantial differences exist between
unicellular and multicellular organisms, but like most cellular
mechanisms, the basic details of homologous recombination appear
conserved throughout all cell types.(15) Decades of genetic,
morphological, and molecular investigations have now established without
question that internal mechanisms are altering the genomes of each cell
used for sexual reproduction, and yet the reactions that are occurring
remain unknown. Despite the absence of specific knowledge regarding the
products of recombination, random mutations are credited for all changes
to gene sequence.
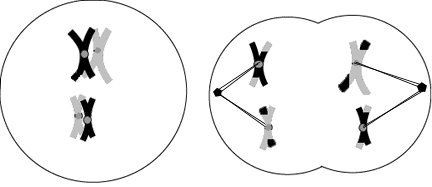 |
Fig 1.
Meiosis
During these stages of
meiosis, homologues pair-up along the cell axis while
recombination takes place, then the genetically unique chromosomes
are pulled apart into daughter cells. Although the exact changes
remain theoretic, it is from these differences that nature
selects, and through which evolution occurs. Another division will
separate the sister chromatids, and generate gametes with only
half of the genetic material possessed
originally. |
During sexual reproduction, an organism is formed
from the fusion of two gametes (egg, sperm, pollen) that are produced
during a unique cell division process called meiosis.(Figure 1) It is
during this portion of the cell life cycle where the parental genomic
DNA is recombined by cellular mechanisms to introduce genetic
variability. The entire nuclear genome condenses during meiosis into
organized units called chromosomes that interact as distinct pairs.
These paired homologous chromosomes possess sequences that are very
similar, and code for variations of the same characteristic like sex
determination. Prior to the division of DNA into new daughter cells, the
homologous pairs of chromosomes are cleaved and spliced together at
multiple locations. During these interactions DNA is exchanged, but the
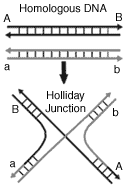
exact details remain unknown. Figure 2 shows a general formation
produced during homologous recombination in prokaryotes and eukaryotes
called a Holliday formation. This splice junction is formed from a
duplex of complementary stands derived from each parental DNA molecule.
The duplex junction has been shown to extend large enough to cover an
entire gene, and ensures exact alignment allowing recombination to occur
under precise control. Many genes involved in homologous recombination
are conserved among all organisms, and in particular, the duplex joints
are formed through reactions mediated by the commonly found
RecA/Rad51-family of proteins.(14)
Fig 2. Holliday
Junction
Recent work has added further
substantiation to the existence of a DNA formation known as the
Holliday junction. During recombination the junction exists as a
series of intermediates during the cascade of reactions involved
with the formation of the recombinant DNA. |
 |
Genes typically exist within a population as a
number of varieties (alleles), and as a result the features they produce
are polymorphic or are present in more than one form. Offspring
variations are largely produced by the intentional recombination of
alleles during meiosis, and it was the regulation of this process that
was demonstrated by Mendel's experiments. Mendelian genetics, and
selective breeding histories have answered a great portion of the
mystery behind heredity. However, the relationship between meiotic
recombination, and the selectable phenotype remains far from
understood.(9) No two offspring are ever the same because the gametes
generated for sexual reproduction are always genetically unique (i.e.
DNA fingerprinting). A relatively short history of recombination, and
artificial selection has led to a tremendous variety of domestic breeds
such as the dogs. Since their discovery, it has been largely assumed
recombination events create these differences by simply rearranging
genes into other combinations, and any new alleles (variations of genes)
are believed to be the result of unintentional mistakes such as
replication errors, or through exposures to external
mutagens.
In addition to sexual reproduction, the
recombination of homologous DNA sequences is induced or shut off as a
programmed cell function during differentiation, development, and is
also stimulated in response to a wide variety of environmental stresses.
It is also well recognized that recombination is controlled by highly
organized regulatory systems.(6) The induction of meiotic recombination
is reliant upon several genes, and is regulated by a complex network of
cellular signaling mechanisms.(14) Furthermore, a cascades of specific
macromolecule interactions has been recognized to coordinate and
regulate catalysis.(10)
Since their discovery or that of genetic mapping it
was assumed that homologous crossing-over during meiosis occurred at
regular intervals along the chromosomes, however, these presumptions
have been largely shattered by a variety of discoveries indicating that
differential recombination rates and patterns exist. For example it is
now well-known recombination frequency is not constant in any one
particular cell. Reactions occur more frequently in some regions of the
genome than in others, with variations of several orders of magnitude
observed. These hyperactive regions have been termed "hot spots" and as
opposed to inert "cold spots" where little to no nucleotide exchange is
found.(3) The process of recombination is likewise recognized to be
significantly different when comparing somatic and germ-line cell types,
and events during meiosis are known to generally occur at a much higher
frequency than in mitosis. For example mitotic recombination frequencies
in the fungus Ustilago maydis have been estimated at 2.9 x 10-7, whereas
in meiosis the rates are closer to 1.9 x 10-3. Sex-specific differences
in recombination frequency are now also relatively well elucidated.
Standard linkage analysis was used to confirm that females have a higher
recombination rate than males, and males recombine preferentially in the
distal regions of the chromosome. These and other techniques were
separately used to establish the existence of significant
inter-individual variation in recombination over short intervals.(13)
Still other researchers demonstrated background effects on the frequency
of recombination using immunostaining techniques to assess meiotic
exchange patterns. It was found in this case and many others that
crossover events were non-randomly distributed and displayed positive
interference.(2)
A New Look from Intelligent Design
Theory
The ability of the cell to intentionally modify the
genome through homologous recombination deserves considerable analysis
from intelligent design theorists. It has become clear from a variety of
sources that indeed organisms possess new genetic information.
Prokaryotes are known to readily uptake polynucleotides, and all major
taxa are able to exchange genes across species barriers through
homologous recombination.(19) It was in fact the genetic transformation
of bacteria in the late 1920's that finally led to the discovery of DNA
as responsible for the heredity of life. Biotechnologists have now also
learned they can easily transform eukaryotes with foreign DNA, and
routinely use this ability to create genetically modified organisms such
as herbicide resistant crops. Many of the procedures used to transfect
plant and animal tissues simply stress the cells or breech the plasma
membrane in the presence of DNA, and some will uptake the genetic
material. (Figure 4) It is now apparent that all cell types can be
genetically transfected with little difficulty, and any extracellular
sequences will be incorporated into the genome if there is sufficient
homology.
 |
Fig. 4 Gene
Gun
Biolistics or the "gene
gun" is the most commonly used technique for performing
genetic transformations. It employs a high pressure delivery
system to shoot tissue with gold or tungsten particles that are
bound by DNA. Although the propulsion is now most typically
accomplished using a gas such as helium, the original variation of
this system from Bio-Rad was equipped with a 22 caliber pistol.
Following a shot with the gene gun, some of the cells will
transport the sequences into the nucleus and splice them into the
genome. |
Genetic samples from living populations have also
shown that closely related species possess an extremely high percentage
of polymorphic DNA. The number of alleles discovered to date is in fact
so high it proves without question they are variations of preexisting
genes. For example the cystathionine ß-synthase gene locus has been
intensely studied in humans due to vascular, skeletal, and nervous
system problems resulting from deficiencies in protein activity. Exon 8
in particular has been found with a high frequency of sequence repeats
or single base variations, and it is estimated that about 5% of human
Caucasians possess alterations in this region.(18) While it is true
random mutations are not likely to accomplish any significantly
advantageous modification, the machinery within the cell has been
overlooked as a source for this new genetic information. The
incorporation of foreign DNA, interspecies gene transfer in bacteria,
and the genetic variations produced during meiosis are all the result of
cellular-performed homologous recombination. These reactions were
intelligently designed to produce these genetic changes, and are widely
known to modify the physical nature of biological organisms through this
process.
Homologous recombination is indeed most important
for its ability to create the offspring variability from which nature
selects, and through which evolution occurs. Following cell division we
cannot determine whether homologous recombination or mutations were
responsible for changes to gene sequence, and yet it has been simply
assumed that any alterations to a reading frame are an unintentional
consequence of these and other DNA manipulations such as replication.
(Figure 5) Environmental adaptations are indeed the result of natural
selection upon individuals with genetic differences, but it is extremely
unlikely that random changes to a complex genetic code could produce any
significant usefulness or evolutionary assistance to an organism.
Despite this rather obvious fact, the theory of evolution requires
random mutations as the ultimate source of new genetic information, and
yet within the last few years the scientific communities best example
has collapsed. The mitochondria were previously thought to offer
evidence that new alleles form through mutation alone because it was
believed the organelle's genome was obtained exclusively from maternal
contributions. However, it has been recently reported in the journal
Science that recombination between parental genomes also occurs in
mitochondrial DNA. Evidence of mixing of paternal with maternal DNA in
the mitochondria was evident, and it was concluded there had been
recombination between the parental genomes.(4,5)
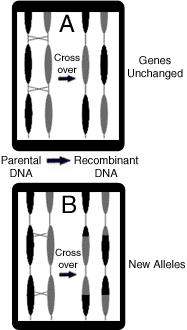 |
Fig 5a. It is believed
the intended function of recombination is to leave existing genes
unchanged by crossing over between reading frames.
Fig 5b.
Crossovers within genes are able to create new alleles, however it
has been assumed any changes to sequence are the result of
unintentional
modifications. |
Although the exchange or movement of alleles from
one homologue to another does occur during meiosis, it is now well
recognized that recombination is not just limited to these procedures.
In addition to creating genetic diversity in offspring, recombination is
involved in error-free DNA repair, and is stimulated by double-stranded
breaks during any stage of the cell cycle. The repair function of
recombination maintains the integrity of the genome through the accurate
correction of several different types of DNA damage.(14) In fact
homologous recombination is now known to be induced by a wide variety of
environmental stresses such as nutrient deprivation, high cell density,
and a great many carcinogens. These reactions are also believed to be
involved with editions such as deletions, duplications, and
translocations between dispersed homologous sequences.(12) The ability
to induce homologous recombination during times of stress would also
provide obvious survival advantages through the introduction of new
genetic information. Mutations would be clearly disruptive in their
affect, however homologous recombination is theoretically able to use
portions of preexisting genes to assemble novel proteins in a systematic
manner.
| "The ability
to induce homologous recombination in response to unfavorable
environmental changes would be adaptive for each species, as it
would increase genetic diversity and would help to avoid
species' extinction. Homologous recombination would be more
efficient for evolution than random mutagenesis or nonhomologous
recombination. Although the latter two will mostly disrupt
previously existing genes rather than creating new ones,
homologous recombination can use previously existing genes as
building blocks, thus enabling the creation of new proteins with
more complex functions in a step-by-step manner. "
Proc. Natl. Acad. Sci. U.S.A.
98(15):8425-8432
(2001) |
Whether mutations are responsible for the
development of any significant useful information is unclear, but the
machinery within the cell cannot be excluded from theoretically
possessing the ability to assemble new gene constructs from portions of
native and foreign sequences. Recently the rapid acquisition of new
functional proteins by homologous recombination was demonstrated in
laboratory experiments, and was shown to be much more efficient than
simple random mutations. Using a technique called DNA shuffling, a pool
of mutated genes was fragmented and reassembled by a self-priming
polymerase chain reaction (PCR). Although not exactly the same as
cellular recombination, it incorporates the formation of a duplex joint
through complementary strands. Using this technique it was demonstrated
that recombination directed the evolution of a gene product with a
significant increase in comparison to simple mutagenesis techniques.
Also, when a mixture of DNA from related organisms was used as starting
DNA, the efficiency of the directed evolution of the gene(s) was
extensively enhanced.(16)
In addition to crossing-over or the exchange of
alleles, there is an entirely new class of homologous recombination only
recently recognized that may share common mechanisms. During the process
of gene conversion, homologous pseudo genes are used to create gene
variations, and are easily distinguished from cross-overs because only
one of the pairs of homologous sequences are altered. During gene
conversion, no other part of the active genome is altered, but instead
regions typically referred to as junk DNA are used in the production of
this variabilty.(Figure 6) DNA repair through conversion is performed
when the regions flanking the damage are replaced by an intact copy from
the sister chromatid or homologous chromosome. It has been documented
that mitotic recombination via gene conversion is able to create
genetically altered cells, and suggested that conversion can generate a
gene with novel functions by rearranging various parts of the parental
open reading frames.(11)
|
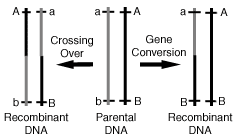 |
Fig 6. Two
Classes of Homologous Recombination
"While meiotic crossing over is supposed to create genetic
diversity by producing new combinations of alleles derived from
parents and the genetic diversity may help cell to adapt to such
unfavorable conditions, the significance of meiotic gene
conversion has not been well understood" Quote and figure reproduced from: Functions
of Homologous DNA Recombination. Takehiko Shibata. Riken
Review 41:21-23 (2001) |
Without a doubt the immunity system provides the
best example of the ability of the cell to generation new genetic
information through homologous recombination. Antibody specificity for
foreign substances is now known to be due primarily to gene editions
that alter the antigen-binding site. It is typically assumed that any
gene variations or alleles are the result of random mutations, and the
changes on the antibody (V-regions) have been largely attributed to such
unintentional alterations. Contrary to these assumptions, the V-regions
on immumoglobulin were first demonstrated to be participating in gene
conversion in chickens, and these reactions are now known to be the
mechanism responsible for V-region alterations in most mammals.(21) The
variable region of the antibody gene is altered through repeated rounds
of intrachromosomal gene conversions that occur between the variable
gene, and a pool of homologous pseudogenes that are being used as donor
sequences.(17) In many vertebrates including humans, the V-region is
altered by a different mechanism that makes single base pair
substitutions instead of through the use of template pseudo genes as
previously mentioned. These alterations have also been assumed to be due
to mutations resulting from relaxed replication fidelity; however, a
clear connection was established this past year between these numerous
individual changes termed "hypermutations", and the process of gene
conversion. A putative RNA editing enzyme (AID - activation-induced
cytidine deaminase) was already known to be required for hypermutations
to occur in the V-region, and has now been recognized as necessary for
gene conversion also. This fact suggests that AID plays a pivotal role
as master controller of antibody gene modifications.(20) Although the
mechanisms of gene conversion within the V-regions remain unknown, these
gene changes are clearly due to cellular mechanisms which produce these
variations with intent.
Conclusion
Current creation science concepts of genetic
heredity are based on early assumptions that are now known to be
incorrect. It has become clear that crossovers do not occur at random
intervals along the chromosome, and likewise that recombination is also
involved with DNA repair and gene conversion. The real function of
recombination during meiosis likely remains misunderstood because
secular science is assigning the tasks they are performing to a source
that is independent of cellular design (mutations). It has been broadly
assumed that genetic variability such as the changes responsible for
antibody specificity are the result of random errors, and yet in most
cases they appear the result of intentional cellular reactions.
Suggestions of randomness in the development of antibody specificity are
also argued strongly by the fact that vaccinations produce certain
outcomes. When a vaccine is effective in a particular species, it will
completely eradicate a disease from the population because every single
individual will develop immunity when inoculated. There are many
examples of viruses such as Polio and Small Pox that have been
completely eliminated from the face of the planet because it is
unquestionable that functional antibodies will be assembled following an
exposure to almost any foreign substance. This acquired immunity is
largely produced through alterations to the genes that produce
antibodies, and random genetic changes cannot with certainty produce a
positive outcome.
The theory of evolution requires an abiotic
mechanism to create new genetic information, and therefore secular
science holds steadfast to Darwin's assumptions despite evidence to the
contrary, and assumes that all new alleles were formed unintentionally.
The undisputed function of these reactions during meiosis is to
recombine parental DNA, and thereby produce offspring variability.
Subsequent selection upon these recombinants has led to the variety of
domestic breeds commonly used by humans today. In nature these changes
provide adaptive capability, therefore the function of genetic
recombination is unquestionably to drive evolution. Although gene
shuffling clearly occurs as a result of this process, it is now well
recognized that foreign or altered genetic codes(alleles) are being used
by various organisms today. Homologous recombination is also now known
to produce genetic diversity through gene conversion, and it is not
illogical to assume the cell is designed to intentionally modify genes
during these reactions. Unfortunately it is largely assumed by most
creation scientists that mutations were responsible for any new alleles.
In fact, a great many creationists do not believe new alleles are
produced as a result of design, and the evolution of groups such as the
canine breeds was simply the result of shuffling preexisting genes.(7,
8)
| "Recombination explains why
children look different from their parents. This shuffling of
the genes can produce superior combinations of different genes.
However, because we see that mutations are incapable of
supplying useful variation, the useful genes that are there to
be shuffled must have been created at the beginning." The
History of Life. Lane P. Lester. Creation Research Society
Quarterly 31(2) 1994 p96 |
However, recent evidence has adequately illustrated
that organisms are uptaking foreign DNA, and using modified native
genes. The percentage of unique alleles found present in closely related
eukaryotes such as humans is certain proof that indeed we are using
altered DNA.(18) We also know all cell types are able to uptake and
incorporate foreign DNA into their genome, and interspecies gene
transfer occurs readily in all major taxa of bacteria through homologous
recombination.(19) There are still further examples of organisms that
appear to have acquired or developed new genetic information such as in
the cases of bacterial antibiotic resistance. In fact many instances
have been found where an organism possesses a unique gene, which gives
it the ability to tolerate an otherwise toxic environment. This fact has
on numerous occasions allowed very important genes to be isolated, and
used for research or medical purposes. For example the bacteria,
(Thermus Aquaticus) was found thriving in Yellowstone hot springs at
temperatures greater than it previously thought possible for life to
exist, and it was therefore recognized to possess proteins that are
stable at higher temperatures than all other cells. The enzymes isolated
from these organisms are now widely used in laboratory reactions
throughout the world for copying polynucleotides using PCR. Such highly
adapted organisms are often found to possess genes not present outside
the habitat in question. This frequent pattern suggests the genes
responsible were either lost in all other related organisms, or were
constructed through a history of recombination while under
regional-specific selection. It is not unreasonable to suggest the
latter may be true at times, and the cell was designed to purposefully
modify genes or create new code to accomplish these evolutionary
adaptations.
The adaptation of an organism to a particular
habitat or niche involves largely uncharacterized modifications of the
genome, and much of what we know about genetic heredity has come from
science that does not recognize the intelligence behind the design of
the cell. Without intelligent design theory, random mutations are the
only other explanation for continued genetic diversity, and are
therefore a theoretic requirement for Darwinian evolution. Despite the
discovery of Mendelian genetics, and recognition that homologous
recombination is responsible for offspring variability, Darwin's
original theories are still taught as fact. Indeed many people today
assume varieties like the finches on the Galapagos are the result of
unintentional changes almost one hundred years following discoveries to
the contrary.
Mutations are a theoretic necessity for the theory
of evolution; therefore, the function of the cell regarding the
production of novel genetic information has been overlooked. The purpose
of genetic recombination is to edit the genome in a largely
uncharacterized manner, and therefore any alterations found to exist are
probably the result of these reactions. This simple logic has escaped
scientists who instead require mutations to be ultimately responsible
for evolutionary adaptations. Creation science perspectives concerning
the existence, and use of new information have also been affected by
erroneous Darwinian assumptions. However, the existence of new alleles
indicates the cell has the ability to generate continued diversity
through recombination, and intelligent design theorists should therefore
closely investigate the ability of the molecular machinery to
purposefully assemble new reading frames from portions of native and
foreign DNA. Although it is clear that many organisms possess and are
utilizing foreign or altered DNA, the participation of the cell in these
processes is unequivocal.
"It is a considerable strain on one's credulity
to assume that finely balanced systems such as certain sense organs
(the eye of vertebrates, or the bird's feather) could be improved by
random mutations. This is even more true of some ecological chain
relationships. However, objectors to random mutations have so far been
unable to advance any alternative explanation that was supported by
substantial evidence". {Harvard biologist
Ernst Mayr, SYSTEMATICS & THE ORIGIN OF SPECIES, 1942,
p.296}
Article References
(1) Evolutionary Biology. Eli Minkoff.
Addison-Wesley Publishing. (1984)
(2) Genetic
control of Mammalian meiotic recombination. I. Variation in exchange
frequencies among males from inbred mouse strains. Koehler KE,
Cherry JP, Lynn A, Hunt PA, Hassold TJ. Genetics 162(1):297-306
(2002)
(3) Meiotic
recombination hotspots. Lichten, M. & Goldman, A.S.H. Annu. Rev.
Genet. 29:423-444 (1995)
(4) mtDNA Shows Signs of Paternal Influence.
Strauss. Science 286:2436 (1999)
(5) Linkage Disequilibrium and Recombination in
Hominid Mitochondrial DNA. Awadalla, Philip; Eyre-Walker, Adam; and
Maynard Smith, John. Science 286:2524-2525 (1999)
(6) Hierarchic
Regulation of Recombination. Kunihiro Ohta. RIKEN Review 41:28-29
(2001)
(7) Genetics No Friend
of Evolution. Lane Lester. Creation Ex Nihilo 20(2):20-22
(1998)
(8) The
History of Life. Lane Lester. Creation Research Society Quarterly
31(2):95-97 (1994)
(9) The Growth of Biological Thought: Diversity,
Evolution, and Inheritance. Ernst Mayr. (1982)
(10) Cascades
of Non-covalent Protein-protein and Protein-DNA Interactions for
Homologous DNA Recombination. Takehiko Shibata. RIKEN Review 46:
(2002)
(11) Functions
of Homologous DNA Recombination. Takehiko Shibata. RIKEN Review
41:21-23 (2001)
(12) Homologous
Recombination as a Mechanism for Genome Rearrangements: Environmental
and Genetic Effects. Alexander Bishop. Human Molecular Genetics
9(16):2427-2434 (2000)
(13) Counting
Cross-overs; Characterizing Meiotic Recombination in Mammals. Terry
Hassold. Human Molecular Genetics 9(16):2409-2419 (2000)
14) Homologous
genetic recombination as an intrinsic dynamic property of a DNA
structure induced by RecA/Rad51-family proteins: a possible advantage of
DNA over RNA as genomic material. Shibata, T., Nishinaka, T.,
Mikawa, T., Aihara, H., Kurumizaka, H., Yokoyama, S. & Ito, Y. Proc.
Natl. Acad. Sci. U.S.A. 98(15) 8425-8432 (2001)
15)
Regulation of meiotic recombination and prophase I progression in
mammals. Cohen P.E. & Pollard J.W. BioEssays 23:996-1009
(2001)
16) Rapid
Evolution of a Protein In-Vitro by DNA Shuffling. Stemmer, W.P.C.
Nature 370(6488):389-391 (1994)
17) The
chicken B cell compartment. Weill JC, Reynaud CA. Science
238(4830):1094-8 (1987)
18) High prevalence of a mutation in the
cystathionine ß-synthase gene. Tsai MY, Bignell M, Schwichtenberg K,
Hanson NQ Am.J.Hum.Genet. 59:1262-1267 (1996)
19) Homologous recombination in procaryotes. Smith,
G.R. Microbiol. Rev. 52:1-28 (1988)
20) AIDing
diversity. Jennifer Bell. Nature Reviews Immunology 2, 223
(2002)
21) Immunoglobulin
diversity, B-cell and antibody repertoire development in large farm
animals. J.E. Butler Rev. sci. tech. Off. int. Epiz., 17(1):43-70
(1998)